

گیرنده هورمون همانطور که از اسم آن مشخص است پروتئین یا مولکلولی است که هورمون برای اینکه بتواند تأثیر خود را اعمال کند باید به گیرندههای خود در سلول هدف متصل شود. گیرندهها معمولاً مولکولهای گلیکوپروتئینی درشت هستند که تعداد آنها از ۲،۰۰۰ تا ۱۰۰،۰۰۰ عدد در یک سلول متغیر است. هر گیرنده، معمولاً برای یک هورمون، واحد اختصاصی است. بدین ترتیب، نوع هورمونی که بر هر بافت معین اثر میکند، مشخص میشود. محل گیرندههای انواع مختلف هورمونها به شرح زیر است:
- درون غشاء یا روی سطح غشاء: مانند هورمونهای پروتئینی، پپتیدی و کاتکول آمینی
- در سیتوپلاسم سلول: مانند هورمونهای استروئیدی
- در هسته سلول و کمپلکس کروموزوم: مانند هورمونهای تیروئیدی
به گیرندههای هورمونهای استروئیدی و تیروئیدی، گیرندههای داخل سلولی گفته میشود. هورمونهای استروئیدی نیز بعد از اتصال به گیرندههای خود در سیتوپلاسم، به هسته سلول منتقل شده و باعث افزایش یا کاهش بیان یک سری از ژنها میشوند.
پیشنهاد میکنیم مقاله هورمون چیست را نیز در سلامتانه مطالعه کنید.
لازم به ذکر است که هورمونهای استروئیدی و تیروئیدی یک سری اثرات غیر ژنومیک نیز دارند. یعنی گیرندههای سطح غشایی هم دارند ولی اثر تیپیک (اصلی) و کلاسیک آنها افزایش یا کاهش بیان یک سری ژنها است.
گیرندهها جز ثابت سلول نیستند. یعنی تعداد آنها میتواند کم و زیاد شود و حساسیت آنها به لیگاند یا ماده شیمیایی نیز متغیر است. پس با تغییر شرایط فیزیولوژیک، تعداد و خصوصیات گیرندهها تغییر میکند.
تعداد گیرندهها در هر سلول هدف، معمولاً از هر روز به روز دیگر یا حتی از دقیقهای به دقیقه دیگر، ثابت نمیمانند. میزان حساسیت گیرندهها نیز توسط مکانیسمهای ویژه تنظیم میشود.
گیرنده غشایی هورمونها
در ادامه میخواهیم ابتدا روی گیرنده های غشایی که سه نوع هستند زوم کنیم. ولی قبل از آن باید مطالبی را در ارتباط با cAMP (یک پیامرسان ثانویه) بلد باشیم.
گیرندههای هورمونی غشایی مرتبط با کانال یونی
به کانال یونیمتصل هستند. عمل مستقیم دارند. از این دست گیرندهها میتوان به گیرندههای نوروترانسمیترها، نوراپینفرین و استیلکولین اشاره کرد.
گیرندههای هورمونی مرتبط با G-پروتئین
این گیرندهها، یک قسمت خارج سلولی، یک قسمت داخل سلولی و یک قسمت میانی دارند. قسمت میانی آن ۷ بار در عرض غشاء عبور کرده است.
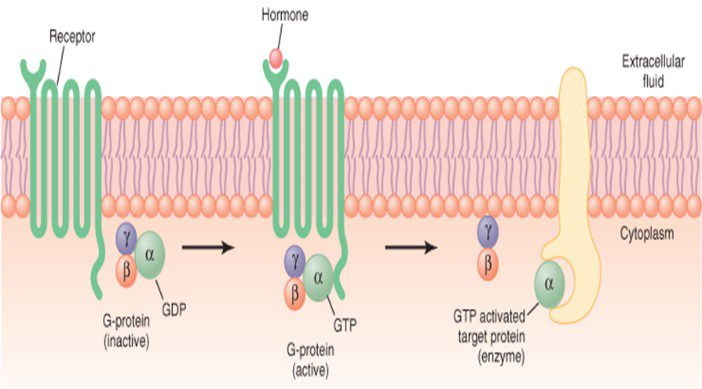
گیرندههای نوع دوم، به واسطهی پروتئینهای متصلشونده به GTP) G-پروتئین) که یک پروتئین واسطه است کار خود را انجام میدهند. به لحاظ ساختاری، G-پروتئینها، سه بخش دارند (β، α و γ) و در واقع تریمر هستند. G-پروتئینِ غیرفعال، در داخل سلول، به سطح داخلی غشاء چسبیده و جزء α متصل به یک GDP (گوانوزین دیفسفات) است.
هنگام اتصال هورمون به گیرندهی خود، گیرنده فعال شده و دچار تغییر شکل فضایی میشود. در این هنگام، گیرنده به G-پروتئینی که خود به GDP متصل است، وصل میشود. سپس GDP از جزء α G-پروتئین جدا شده و به جای آن یک GTP به جزء α متصل میشود. این رویداد، سبب فعال شدن G-پروتئین شده و جزء α همراه با GTP متصل به آن، از G-پروتئین جدا میگردد و به آنزیمی در غشای سلول متصل میشود و آنزیم را فعال میکند. به این آنزیم، آدنیلیلسیکلاز میگویند. (برای درک بهتر، خط به خط مراحل را با شکل تطبیق دهید)
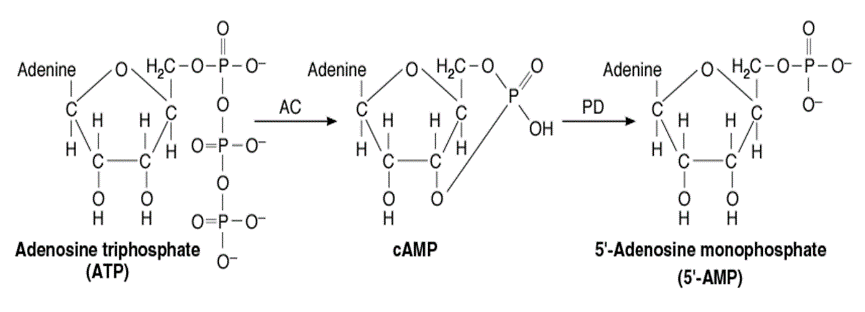
به دنبال فعال شدن آنزیم آدنیلیل سیکلاز، ATP به cAMP تبدیل میشود. همان طور که گفته شد cAMP نیز نقش پیک ثانویه را دارد. cAMP یک سری پروتئینکینازهای وابسته به cAMP را که در سلولها به صورت غیرفعال هستند، به شکل فعال درمیآورد. پروتئینکینازها پروتئینهایی هستند که یک گروه فسفات (PO43-) به مادهی اولیه اضافه میکنند. وقتی پروتئینکینازها فعال شدند، گروه فسفات را به یک سری از پروتئینها در داخل سلول اضافه میکنند. از جمله این پروتئینها، آنزیمها هستند که فسفریله شده و باعث ایجاد پاسخ سلولی میشوند.
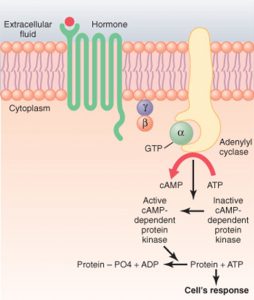
داستان را که دنبال میکنید؟! خب! گفتیم که پس از فعال شدن G-پروتئین و زیرواحد α، این زیرواحد α از G-پروتئین جدا میگردد و با اتصال به آدنیلیلسیکلاز در غشای سلول، آن را فعال میکند. حالا برویم و داستان را ادامه بدهیم!
بعد از فعال شدن آدنیلیل سیکلاز، جزء α مجدداً غیر فعال شده و GTP جای خود را به GDP میدهد و عمل پیامرسانی به سرعت متوقف میشود. سپس بار دیگر زیرواحد α به زیرواحدهای β و γ متصل میشود و همان G-پروتئین اولیه و غیرفعال را تشکیل میدهد.
گفتیم در صورت اتصال هورمون به گیرندهی خود، G-پروتئین فعال میشود. حالا G-پروتئین فعال، میتواند اثر تحریکی یا مهاری داشته باشد. در صورتی که از علامت اختصاری «Gs» استفاده شود، منظور نقش تحریکی آن و در صورت استفاده از «Gi»، منظور نقش مهاری آن است. به عبارت دیگر، یا فعالیت آنزیم آدنیلیل سیکلاز افزایش مییابد (Gs) و یا از فعالیت این آنزیم کاسته میشود (Gi).
در صورتی که G-پروتئین نقش تحریکی داشته باشد، آدنیلیل سیکلاز باعث تولید cAMP میشود و cAMP نیز باعث فعال شدن پروتئینکینازهای وابسته به cAMP میگردد و آنها را به فرم فعال تبدیل میکند.
پروتئینکینازها روی یک سری آنزیمها، گروه فسفات اضافه کرده و فعالیتهای بیولوژیکی به صورت آبشاری انجام میشود. همچنین پروتئینکیناز فعال میتواند بر روی DNA هسته نیز اثر گذاشته و باعث افزایش یا کاهش بیان یک سری ژنها شود.


گیرنده های هورمونی مرتبط با آنزیم
گیرندههای هورمونی مرتبط با آنزیم، وابسته به G-پروتئین نیستند و خودشان به عنوان آنزیم عمل میکنند یا آنزیمهایی را که در ارتباط نزدیک با گیرنده هستند، فعال میکنند. ساختمان آنها متفاوت با انواع وابسته به G-پروتئین است. این گیرندهها صرفاً یک بار از عرض غشاء عبور میکنند (نه ۷ بار) و دارای یک قسمت خارج سلولی، یک قسمت که از غشاء عبور میکند و یک قسمت داخل سلولی هستند. قسمت داخل سلولی، پایانه C نام دارد.
همانطور که در شکل دیده میشود، گیرندههای دستهسوم، دو نوع میباشند:
❶گیرندهای که در سمت چپ شکل قرار دارد صرفاً یک بار از غشاء عبور کرده و یک بخش خارج سلولی و یک بخش داخل سلولی دارد. وقتی هورمون به گیرنده وصل شد، در قسمت داخل سلولیِ آن، فرایند اتوفسفوریلاسیون صورت میگیرد و باعث میشود که به صورت تیروزینکیناز فعال درآید. تیروزینکیناز فعال سبب اضافه شدن مولکولهای پروتئینی شده و باعث انجام یک سری فعالیتها میشود از جمله ۱- جابهجایی حاملها، ۲- فعال کردن آنزیمهای دیگر و ۳- فعال کردن فاکتورهای نسخهبرداری و تأثیر در بیان ژنها. نمونهای از این نوع گیرندهها، گیرندهی هورمون انسولین میباشد.
❷در سمت راست شکل، نوع دوم گیرندههای دستهی سوم نمایش داده شده است. گیرندههای نوع دوم، از لحاظ ساختمانی مثل نوع اول میباشند. وقتی هورمون به گیرنده متصل میشود، تغییراتی در شکل فضایی گیرنده رخ میدهد و یک سری آنزیمهایی که در ارتباط با بخش داخل سلولی هستند فعال شده و فسفوریله میشوند. به ویژه وقتی روی آمینواسید تیروزین فعال میشوند، میتوانند یک سری فاکتورهای نسخهبرداری را از طریق فسفریلاسیون فعال کنند و این امر باعث افزایش یا کاهش بیان یک سری از ژنها میشود. نمونهای از این گیرندهها، گیرنده هورمون رشد (سوماتوتروپین) و گیرنده لپتین (هورمون تنظیم انرژی و اشتها) است.
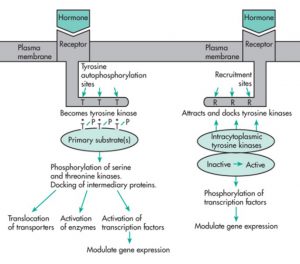
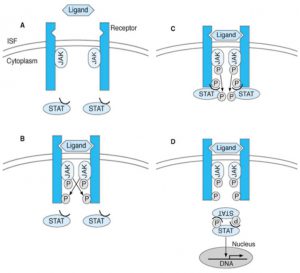
لپتین را به عنوان هورمونِ لیگاند در نظر میگیریم که گیرندهی آن به صورت دایمر (دوتایی) است. وقتی لیگاند به گیرنده متصل میشود، آنزیمهای تیروزین کیناز به نام JAK که در ارتباط نزدیک با بخش داخل سلولیِ گیرنده هستند و نیز بخشهای داخل سلولی گیرنده، فسفریله میشود. وقتی کمپلکس فعال شد، فاکتورهای نسخهبرداری به نام STAT (پروتئینهای تبدیلکنندهی پیام و فعالکننده رونویسی) را فسفریله و فعال کرده و دایمری تشکیل میدهند که به DNA متصل شده و باعث ۱- افزایش بیان یک سری از ژنها و ۲- فعال شدن مسیرهای آنزیمی دیگر مانند MAP و PI3K میشوند.
گیرندههای داخل سلولی هورمون ها
گیرندههای داخل سلولی در مورد هورمونهای استروئیدی و تیروئیدی مطرح هستند. قبلاً هم گفتیم که گیرندهی هورمونهای استروئیدی در سیتوپلاسم سلول و گیرنده هورمونهای تیروئیدی در داخل هسته و در کمپلکس کروموزومی قرار دارد.
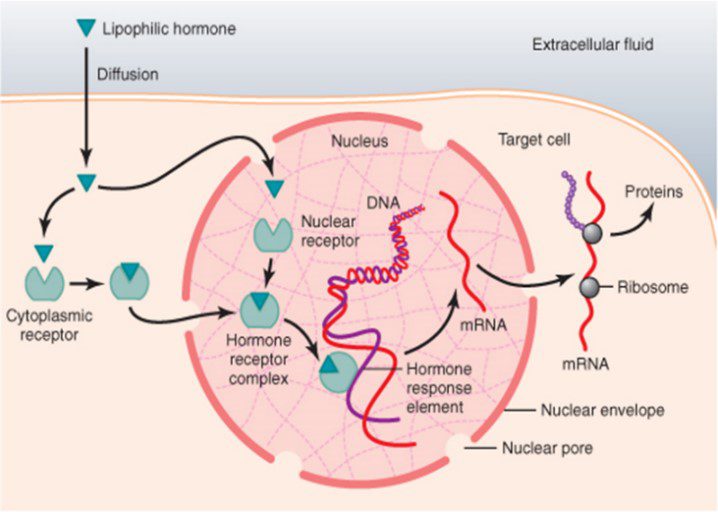
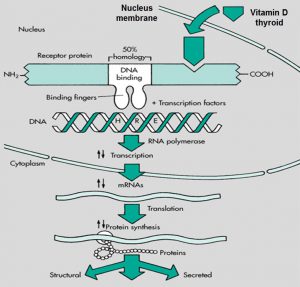
هورمون استروئیدی بنام Lipophilic hormone در شکل نشان داده شده که به خاطر ساختمان استروئیدی خود به راحتی از غشاء عبور کرده و به گیرندهی خود در سیتوپلاسم متصل میشود و سپس این کمپلکس از سیتوپلاسم به سمت هسته رفته و به قسمتی در DNA تحت عنوان جزء پاسخدهنده به هورمون (HRE)، متصل میشود. این قسمت، ناحیهی پروموتور است و باعث افزایش یا کاهش بیان یک سری از ژنها میشود و از این طریق فعالیت خود را انجام میدهد.
گیرندهی هورمون تیروئیدی وقتی به ناحیهی HRE در DNA متصل میشود، تأثیر خود را به صورت افزایش یا کاهش بیان ژنها اعمال میکند.
بعضی از اعمال هورمونهای استروئیدی سریع است که با مکانیسم عمل آنها (مکانیسمی که از طریق هسته فعالیت خود را انجام میدهد) در تضاد است. محققان متوجه شدند که یک سری گیرندههای غشایی نیز برای هورمونهای استروئیدی مطرح هستند که باعث تولید پیامبر ثانویه میشوند. این عمل که به وسیله گیرندههای غشایی انجام میشود، اعمال غیرژنومی نامیده میشوند.
فعالیت از طریق پیک ثانویه هورمون ها
هورمونهای پپتیدی و کاتکول آمینی، پیک اولیه هستند که با اتصال به گیرندههای روی سلول هدف، پیک ثانویه ایجاد میشود. پیک ثانویه ادامه دهندهی فعالیت هورمون در داخل سلول است.
چند پیک ثانویه معروف که در ادامه به تفکیک بررسی خواهیم کرد عبارتند از:
- cAMP (آدنوزین مونوفسفات حلقوی): توسط آدنیل سیکلاز (آدنیلیل سیکلاز) بدست میآید.
- کلسیم-کالمودولین: کالمودولین یک پروتئین است که در عملکرد و ساختار خود شبیه به تروپونین C است.
- فسفولیپیدهای غشای سلولی: در نتیجه تجزیه فسفولیپیدهای غشای سلول ایجاد میشوند.
cAMP (آدنوزین مونوفسفات حلقوی)
در ارتباط با پیک ثانویه cAMP-آدنیلیل سیکلاز، در شکل های زیر به طور مفصل صحبت کردیم. ولی به طور خیلی خلاصه دوباره میگیم بهتون!
هورمون به گیرنده متصل به G-پروتئین وصل شده و آنزیم غشایی آدنیلیل سیکلاز را فعال میکند. این آنزیم ATP را به cAMP تبدیل کرده که در ادامه آنزیم دوم یعنی PKA (پروتئینکیناز وابسته به cAMP از نوع A) فعال شده و بقیه پروتئینهای لازم را فسفریله میکند و آبشاری از واکنشها را به راه میاندازد.
سیستم پیک ثانویه کلسیم-کالمودولین
در این مورد، G-پروتئین باعث باز شدن کانال کلسیم شده (نقش واسطه) و هنگام باز شدن کانال، Ca2+ طبق شیب غلظت، وارد سلول میشود. البته این کانال یونی متفاوت از گیرندههای دستهی اول (❶) است که قبلاً مطرح شد.
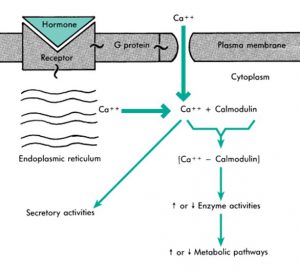
وقتی Ca2+ به داخل سلول حرکت میکند غلظت Ca2+ در سلول زیاد میشود. Ca2+ به پروتئین کالمودولین متصل میشود (کالمودولین گیرنده Ca2+ است) و یک کمپلکس ایجاد میشود که
فعالیت یک سری آنزیمها را افزایش یا کاهش میدهد و مسیر یک سری فعالیتهای متابولیکی را فعال یا غیرفعال میکند. پس در این مسیر، G-پروتئین یک پیک ثانویهی دیگری را تحت عنوان کلسیم-کالمودولین، فعال میکند.
ساختار کالمودولین نشان میدهد که میتواند به چهار یون Ca2+ متصل شود. وقتی با ۴ یا ۳ یون کلسیم band شود (همون وصل شود!) میتواند چندین نوع مختلف از کینازهای وابسته به کالمودولین را فعال کند، زیرا همانطور که گفته شد کمپلکس کلسیم-کالمودولین، یک پیامبر ثانویه است.
از جمله کینازهای وابسته به کالمودولین، میتوان به سه مورد اشاره کرد: ۱- کیناز زنجیره سبک میوزین (که مستقیماً روی میوزین عضلهی صاف عمل کرده و سبب انقباض عضلهی صاف میشود)، ۲- فسفریلاز کیناز و ۳- کلسیم-کالمودولین کیناز I، II و III (که هر کدام فعالیت متفاوتی دارند).
یون Ca2+ تعداد زیادی از فرایندهای فیزیولوژیک را تنظیم میکند. از جمله فرایندهای رشد، سیگنالرسانی عصبی، یادگیری، انقباض، ترشح و لقاح. تنظیم غلظت کلسیمِ داخل سلولی بسیار مهم است.
فسفولیپیدهای غشای سلولی
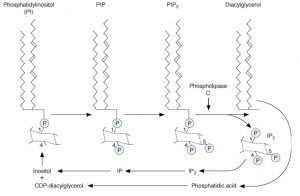
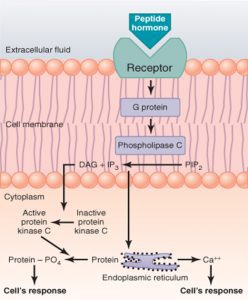
با اتصال هورمون پپتیدی به گیرندهی خود روی غشای سلول هدف، G-پروتئین فعال شده و آنزیمی که در این مسیر فعال میشود، فسفولیپاز C است که متفاوت با آنزیمهای دیگر است. این آنزیم همانطور که از نامش مشخص است، روی فسفولیپیدهای غشاء اثر گذاشته و باعث تبدیل فسفاتیدیل اینوزیتول دیفسفات (PIP2) به دو واسطهی دیآسیل گلیسرول (DAG) و اینوزیتول تریفسفات (IP3) میشود.
IP3 یک پیامبر ثانویه است و باعث افزایش فراخوانی Ca2+ از شبکه اندوپلاسمی به داخل سلول شده و میزان غلظت Ca2+ داخل سلول افزایش مییابد. همچنین IP3 میتواند روی پروتئینکیناز C که غیرفعال است اثر گذاشته و آن را به حالت فعال تبدیل کند. (پس ۲ کار انجام میده). پروتئین کیناز C نیز روی یک سری پروتئینهای داخل سلول اثر گذاشته و با افزودن فسفات به آنها، باعث ایجاد پاسخ بیولوژیک میشود.
وقتی IP3 و DAG کار خود را انجام دادند، مجدداً با هم ترکیب شده و فسفاتیدیل اینوزیتول (PI) را که پیشساز تشکیل PIP2 است، تشکیل میدهند.
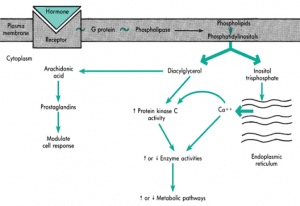
بخش لیپیدی DAG آراشیدونیک اسید است که دستهی چهارم هورمونها تحت عنوان پروستونوئیدها را ایجاد میکند. دستهای از این هورمونها پروستاگلاندین میباشد و به این ترتیب فعالیت سلول را تغییر میدهد.
همچنین DAG، پروتئینکیناز C را فعال میکند و با فسفریلاسیون سایر پروتئینها، یک آبشار واکنشی راه میاندازد.
نقشه راه را که همچنان دارید؟! خب! ما ۳ تا پیک ثانویه معروف را بررسی کردیم. یک پیک ثانویه غیر معروف(!) هم وجود دارد که میخواهیم آن را نیز بررسی کنیم. با ما باشید!
نحوهی ایجاد و عملکرد cGMP
همانطور که در شکل دیده میشود، دو سلول مختلف نمایش داده شده است که سلول سمت راست سلول اندوتلیال عروق و سلول سمت چپ سلول عضلهی صاف عروق است.
وقتی در سلولهای اندوتلیال، نیتریک اکساید (NO) تحت تأثیر NOسنتاز ایجاد شود (در اثر شکستن آرژنین و تبدیل آن به NO)، به راحتی از غشای سلول عبور کرده (چون NO یک گاز است) و از سلول اندوتلیال وارد سلول عضلهی صاف میشود و این امر آنزیم گوانیل سیکلاز را که یک آنزیم در داخل سیتوپلاسم است، فعال میکند و GTP به cGMP تبدیل میشود. cGMP یک پیامبر ثانویه است و هنگامی که ایجاد شود، پروتئینکیناز G را فعال کرده و پروتئینکیناز G از طریق مکانیسمی باعث کاهش Ca2+ و شل شدن عضله میشود و به دنبال آن قطر رگ افزایش مییابد.
یلام ممنون از مطالب خوبتون و اینکه انقدر واضح توضیح دادید
خوب و مفید ، ممنون از شما
سلام
بسیار مفید بود از زحمات شما بسیار متشکرم امیدوارم مقالات بیشتری از شما ببینم
موفق باشید
بسیار عالی و مفید بود ممنونم 🙏🏼🌸🌸
موفق باشید